

BY DAVID EYLES
In the middle of all the arguments and to-ing and fro-ing of the debate upon how we should manage our uplands, the issue of whether shooting does or does not have a serendipitous conservation value is once again raging – just as it does every year with the opening of the grouse season on the 12th August, then game shooting and wildfowling today (the 1st September). At my level of consciousness (limited, it is true) the first blows were struck by Peter Glenser, Chairman of the BASC, in this article in the Country Squire Magazine. The right of reply was given to Dr Steve Carver of Leeds University here. There was a further piece written by Liam Stokes and more comments by Steve Carver at the end of Liam’s article.
1.0 – Symbiosis.
There were two things about Steve Carver’s response which caught my eye. The first was his use of the term ‘symbiosis’ to describe the relationship between the predators who interfere with numbers of the game species such as pheasants and grouse. Symbiosis is the term ecologists use to describe two species who live in close proximity and whose habits are in some way mutually beneficial. It is a bit more complicated than that, so I quote verbatim the opening paragraphs from Chapter 13 of ‘Ecology – from Individuals to Ecosystems’ [1].
“No species live in isolation, but often the the association with another species is especially close: for many organisms, the habitat they occupy is that of another species. Parasites live within the body cavities or even the cells of their hosts; nitrogen fixing bacteria live in nodules on the roots of leguminous plants; and so on. Symbiosis (‘living together’) is the term that has been coined for such close physical associations between species, in which a ‘symbiont’ occupies a habitat provided by a ‘host’.
In fact, parasites are usually excluded from the category of symbionts, which is reserved instead for for interactions where there is at least the suggestion of ‘mutualism’. A mutualistic relationship is simply one in which organisms of different species interact to their mutual benefit. It usually involves the direct exchange of goods or services (e.g. food, defense or transport) and typically results in the acquisition of novel capabilities by at least one partner. Mutualism, therefore, need not involve close physical association; mutualists need not be symbionts. For example, many plants gain dispersal of their seeds by offering by offering a reward to birds or mammals in the form of fleshy edible fruits, and many plants assure effective pollination by offering of nectar to visiting insects. These are mutualistic interactions but they are not symbioses.
It would be wrong however, to see mutualistic interactions simply as conflict-free relationships from which nothing but good things flow for both partners. Rather, current evolutionary thinking views mutualisms as cases of reciprocal exploitation where, none the less, each partner is a net beneficiary.”
So that’s it then: symbiosis is where there is a close physical association and a net benefit to both species. So when it comes to considering game species and their predators, we cannot say that it is a symbiotic association because there is no net benefit for both predator and prey. Consider a Peregrine Falcon which kills and eats a pigeon: whilst the Peregrine gains food, there is manifestly no advantage to the pigeon. Neither do the two species live in particularly close proximity – for obvious reasons. In fact ecologists, with admirable simplicity, term the association between predator and prey as a ‘predator-prey relationship’. They may get technical and further describe it as ‘density dependent’ or ‘density independent’, or even ‘functionally dependent’; but we have little need to worry about these nuances here.
I needed to clear this up before proceeding too far because Steve Carver, in introducing the term, has confused his own argument.
2.0 – Predator-Prey Relationships
The second point which caught my eye was the following argument, which is the crux of Steve’s case and which I quote below:
“…predator populations follow in close synchronisation, sometimes a year or two behind their prey species, depending on birth rates and populations. [See Figure 1 below] Thereby land management for shootable surpluses of game species also increases populations of predators such as mustelids, corvids, raptors, foxes etc….. Heavy handed predator control just creates a predator vacuum into which other predators flow, once the a territory is seen as vacant. This further perpetuates for yet more predator control.
To make matters worse, not only do these predators prey on the same species you are trying to protect in order to create the shootable surplus, but there will also be a “spill over” predation on other species of conservation importance such as ground nesting birds (curlew, plover, lapwing) that your predator control also purports to protect, since predator species don’t distinguish between game and non-game birds.”
Figure 1 – Typical theoretical representation of population densities of predators and prey (Source unknown).

This quotation from Steve Carver can be boiled down to a succession of arguments:
- Predator numbers depend upon the density of prey and fluctuate with a lag, of varying time, behind prey numbers (Figure 1).
- In game or grouse shooting, where numbers of prey species are inflated to very high densities, this will ‘drag in’ predators from outside and thereby increase the density of predators.
- When predator control is undertaken, this will create a ‘predator vacuum’ and so draw in more predators.
- Given high densities of predators, this will increase incidental predation upon species of high conservation value.
Figure 1 above is a theoretical model, the actual picture often becoming a lot more confused over time, as indicated by Figure 1 here. However, as a rough guide, it’s not a bad one. This suggests that the first of Steve’s hypotheses stacks up – because it fits in with theoretical and observational studies.
The second of Steve’s hypotheses suggests that the numbers of predators will also rise to much higher levels than normal, because the prey species are at abnormally high densities. This is open to some debate because, in the context of UK game shooting, the size of shooting estates is quite small and will support only a limited number of predators, because the size of the predator territories is a limiting factor. Additionally, the availability of surplus prey is limited over time, i.e. there is a very short-term glut of prey species and a limited number of predators, meaning that the impact of the predators is relatively small. This is in some ways analogous to the situation in the Arctic tundra where sudden peaks of voles and lemmings overwhelms the resources of the predators in the area to take out a given proportion of the prey. These objections aside, in theory hypothesis number two might stack up reasonably well.
The third hypothesis is common-sense, except insofar as there is a time lag between getting rid of the predators and their return from outside. The rapidity with which predators re-populate the area depends upon the species – avian species are generally quicker to return because they are more mobile. Furthermore, they will tend to be younger individuals because these are the ones which disperse from their home ranges. It will often take a year or two for a young raptor to settle in and find a mate before it begins to breed and so exploit the prey in the area. This timescale of a year or two needs to be contrasted with the very short window of glut (i.e. seasonal) by the management of game birds.
The fourth hypothesis is a product the generalist nature of most predators, who are able to switch from their preferred species to another, on an opportunist principle. A Common Buzzard, who might normally specialise in grouse chicks and voles on a grouse moor, will not pass up the opportunity of a Curlew chick, if one becomes available.
3.0 – Wolves, Moose and Caribou.
So far, Steve Carver’s reply to Peter Glenser seems to hold up reasonably well. But a week or two after these responses were published by Country Squire Magazine, a paper was published from a Canadian study of the effects of controlling Moose in order to reduce the Wolf predation upon Caribou. The New York Times article which alerted us to the publication is here and the paper itself is here. Steve Carver’s response was to suggest that the Canadian management strategy undertaken by this experiment could be an example for UK game and grouse shooting to follow (and, by implication, simultaneously allowing rewilding into the frame).
The species of particular interest in this paper[2] is the North American Caribou, which is a group of subspecies of the Eurasian Reindeer Rangifer tarandus. There are four subspecies of Caribou; and the particular subspecies under scrutiny by this study is the Woodland Caribou Rangifer tarandus caribou which, as its name suggests, lives primarily in the boreal forests of Canada. Caribou have declined dramatically in recent years and so are of considerable conservation interest. One reason for their decline is given as the increase in numbers of Moose Alces alces and to a much lesser extent, White-Tailed Deer Odocoileus virginianus. These species have moved into the forests of Canada in much greater numbers because large scale logging activities have altered the mature forest into a mix of vegetation which favours the Moose and White-Tailed Deer, but also reduces the amount of arboreal lichens which is the main winter food of the Caribou. Along with the Moose come large increases in Wolves Canis lupus whose primary prey is the Moose, but which will also take Caribou.
The ecological principle around which this study hinges is that of apparent competition. This relies upon the idea that when a novel prey species colonises an area, predators arrive with it. The predators then also suppress secondary, but native species. If the native species are unable to reproduce fast enough to replace their losses, this will put them on a downward path to extinction or local extirpation. In this case, the native species is the Caribou, the novel species is the Moose (and White-Tailed Deer) and the predator, now arrived in big numbers, is the Wolf. The central hypothesis of this study is that a large reduction of Moose numbers to an ecologically reasonable target (about a 70% reduction), will reduce wolf numbers sufficiently to assist the Caribou numbers to recover. Within this general hypothesis are three specific hypotheses, each depending upon the preceding one:
- Sport hunting can be used to reduce the numbers of Moose.
- Wolves are primarily limited by Moose abundance.
- Caribou are primarily limited by Wolf abundance.
The study set out to test these hypotheses by dividing the study area of two mountain ranges within British Columbia into a treatment area and a reference (control) area. The treatment area was 6,500 km² and the reference area was 11,500 km². Within these areas, there were a total of five named sub-populations of Caribou of widely varying sizes. The numbers of Moose that were taken out was about 535 from the treatment area over the years of 2003 to 2005, whilst the numbers taken out from the reference area was about a dozen a year, which was usual practice. Wolf abundance was monitored using GPS and VHF collars on chosen individuals.
Figure 2 – Number of Moose harvested in treatment and reference areas (Source: screenshot from Serrouya et al. 2017).

Note that the numbers of Moose culled in the treatment area after 2005, continues to be higher than in the reference area. This is necessary to maintain Moose abundance at the desired levels, otherwise reproduction will simply build the numbers up again.
As would be expected from the theoretical model of predator-prey relationships, the Wolves declined in response to the reduced numbers of their primary prey species, which were achieved after the concentrated culling during the years 2003 to 2005:
Figure 3 – Moose and Wolf abundance in the treatment areas from 2002 to 2014. Note that wolf abundance monitoring was started in 2007. (Source: Serrouya et al. 2017).
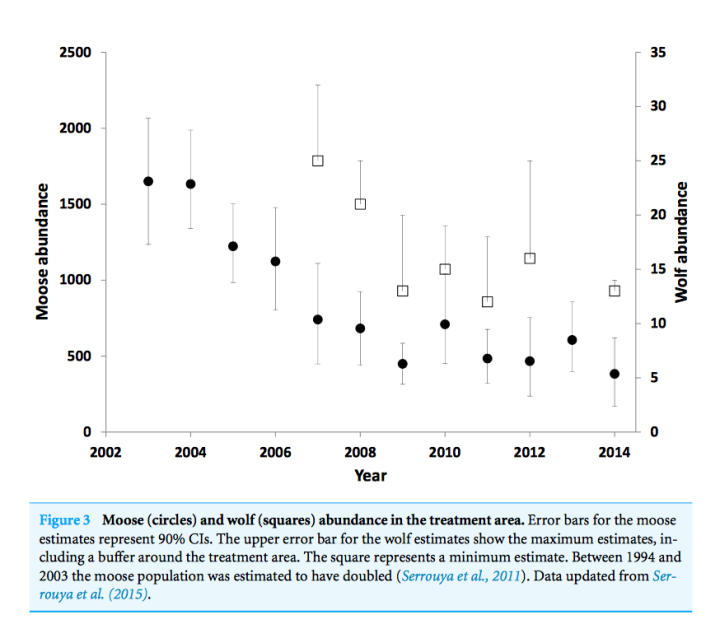
Note that Moose culling started in 2003, but monitoring of Wolf abundance only started in 2007, so there is a gap in information of four years as to what was happening to the Wolves. Although it is not possible to determine exactly when the Wolves began to decline in response to the reduced number of Moose, it is clear that a decline happened and continued until the Moose abundance levelled out from 2009 onwards.
To test the effectiveness of severe culling of Moose upon the populations of Caribou, the abundance of each sub-population of caribou was recorded before and after treatment began in 2003.
Figure 4 – Caribou abundance estimates, before and after treatment began (dotted line), for each sub-population (Source: Serrouya et al. 2017).

In Figure 4, the top three graphs represent the fluctuations in Caribou abundance for the three treatment sub-populations. The bottom two are the same for the reference sub-populations. The Columbia North (treatment) and Wells Gray South (reference) are the largest sub-populations, each occupying about 20% of the total study area. The two smallest areas are Frisby-Queest (treatment) and Groundhog (reference). Columbia South (treatment) is intermediate in size.
These graphs show that in the small and intermediate areas, Caribou numbers continued to decline dramatically after 2003, regardless of whether or not those areas had been subjected to treatment. The largest reference area (Wells Gray South) suggests that Caribou numbers continue to decline, but more slowly than in the smaller areas. Only Columbia North (treatment) shows a stabilisation of Caribou numbers, with the possibility of a small increase by 2014. As Caribou have a low rate of reproduction recovery from a high risk of predation to a much lower risk will clearly take some years. It could be argued that Caribou numbers will recover further but slowly. But from the paper, this cannot be said to be absolutely certain. The smaller areas and sub-populations are perhaps more interesting in that they suggest that lurking confounders are at play – such as food availability declines.
Overall, the first two of the hypotheses listed above appear to be supported: i.e. that Moose numbers can be quickly reduced by sport hunting; and that Wolf numbers are limited by the abundance of their principal prey species (i.e. Moose). The third hypothesis that Caribou are primarily limited by Wolf abundance is much less well supported. Despite the apparent confidence of the title of the paper, the data suggest that caution (and a good deal more work) is needed before the success of the treatment can be announced with complete confidence.
4.0 – Are Wolves, Moose and Caribou relevant to British Grouse Moors?
Outwardly, in the sense that Steve Carver means it, there are a number of beguiling similarities between the Woodland Caribou in the boreal rainforests of British Columbia, and the management of a large sporting estate in England or a grouse moor in Scotland. These similarities can be listed as follows:
- The Canadian Moose can be thought of in the same way as a pheasant on our hypothetical sporting estate. Both are introduced interlopers upon the existing natural environment.
- The Wolf, as a generalist predator (albeit one with particular preferred tastes), has its parallels in the Fox, Common Buzzard, Carrion Crow, Magpies and so on.
- The Caribou, as a native species with a low reproductive success, has its parallels in the form of (say) a Lapwing which mimics the low success of the Caribou because its reproductive strategy is intrinsically high risk.
- Caribou numbers appear to be limited by Wolves (but other factors may also contribute); Lapwing are limited by foxes and corvids (but are also limited by modern agricultural methods).
- Both Caribou and Lapwing are declining rapidly and both are of considerable importance from a conservation point of view.
- Both the Canadian study and the UK sporting estate operate strategies of continuous management. The Moose continue to be culled because of reproductive replacement. In the UK, predators are also continuously controlled both because of reproductive replacement and also inward migration from outside the area.
These are the similarities which come to mind. However, there are also differences:
- The most obvious disparity is that of scale. Most UK sporting estates, even large ones in Scotland are a matter of a few hundred hectares at the most. The area for this Canadian study is 18,000 km² or roughly 87% the size of the whole of Wales. Its size and extremely rugged landscape is why monitoring of the radio tagged Wolves had to use a fixed wing aircraft in order to cover the vast area adequately. Size really does matter. The bigger the contiguous habitat, the lower the proportion of edge effect. This edge effect is important because the size and physical terrain limited the flow of Wolves and Moose into and out of the Canadian study area. This becomes even more important when there are hard, physical boundaries separating areas of interest and which limit or stop the movement of predators. There is a particularly rugged mountain range (Monashee Mountains – see Figure 5) to divide the treatment area from the reference area. Inevitably, this sort of physical barrier will slow up the immigration/emigration of large mammals in a way that does not happen in the UK – where many of the predators are fast moving, often avian and the landscape is nothing like as rugged as in British Columbia.
- The Canadian study had large mammals as its subjects. Population responses to increases in predation, and then withdrawal of that predation, tend to be measured over years. A sporting estate in the UK has a very brief surplus for little more than two or three months. This may allow those resident predators to gorge themselves for a happy, but ephemeral period; after that, prey populations return to normal levels very quickly. Increased numbers of predators moving into such an area to enjoy the surplus will quickly find a very lean period after the 31st January at the end of the shooting season. This inhibits the build up of a large number of predators during the breeding season that follows.
- The Canadian study was pursuing a path of indirect control: cut the numbers of Moose to reduce the numbers of Wolves, which in turn helps the Caribou. In UK sporting estates and smaller shoots, the approach is much more direct: reduce the number of predators to reduce the effect upon both the game species and the native species of conservation interest, such as Lapwing or Curlew.
- Perhaps the principal difference is that of the objectives between the Canadian study and sporting estates in the UK. The Canadian study had two problem groups which it wanted to reduce: the novel species (Moose) and the predator species (Wolves). A UK sporting estate has only one problem group: that of the predators (which, although they are more than one species, for the purposes of this argument can be considered as one problem). Conversely, the Canadian study had one native species as its conservation objective (Caribou): whereas the UK situation has two conservation objectives (Pheasant or Grouse, and Lapwing, Curlew etc).
- The UK sporting interest provides a huge financial benefit to the landowners and farmers. Much of that money trickles down into the local economy in the form of wages and the service economy. In addition, a great deal more money comes into the local economy because of tourism which is prompted by the landscape itself, managed jointly by farmers and the sporting interests. The amount of money that flows into the area in this way, far exceeds (on a per hectare basis) any equivalent amounts generated by the British Columbia landscape, magnificent though it is. In particular, the Canadian sporting interest will generate a tiny amount per hectare compared with the UK situation.
Figure 5 – Monashee Mountains separating the treatment from the reference areas (Source: Google images).

Conclusions
Steve Carver’s overall thrust seems to be that if UK sporting interests (and farmers) were to accept lower numbers of game for a shootable surplus (or lower yields), this would enable a ‘rewilded’ landscape to ‘improve’ ‘biodiversity’ and ‘wildness’. These claims are open to criticism and are unlikely to prove viable, for at least some of the points given above.
Regardless of ecological theory, or experiments conducted on a vast scale in Canada, the simple fact of the matter is that direct predator control as practiced in Britain, succeeds in protecting both game species and species of high conservation interest. The daily experience of every gamekeeper, and which gets them out of bed early in the mornings, is now backed up by a growing body of scientific evidence which shows that land managed for sporting purposes supports a higher diversity of species of conservation interest than similar land. Over decades of work by the late Dick Potts[3] and colleagues at the Game and Wildlife Conservation Trust, has repeatedly demonstrated that the management and creation of particular habitats (such as beetle banks in arable fields), as well as predator control, will greatly increase numbers of birds and other species. Predator control is now conducted routinely by the RSPB on some of their reserves, and they are also successful advocates of eradication of rats, mice and hedgehogs from naive island populations of breeding seabirds. Studies on predation of wader species are discussed by Ian Newton[4] who says: “Once predators have been removed from an area, nest success typically shows a substantial increase, and in subsequent years breeding densities rise…..Moreover, in the uplands, waders show greater nest success and nesting densities in areas where predators are controlled by gamekeepers than in other similar habitat elsewhere.” These statements are again backed up by a very recent paper on Curlew Numenius arquata breeding abundance by Franks et al.[5]. This statistical modelling study examined the mapped records provided by the successive breeding bird surveys organised by the British Trust for Ornithology. They used the Breeding Bird Surveys from 1995 and 2011 at a resolution of 1km² to investigate the potential drivers of variation in curlew population change across Great Britain. Under the section headed ‘Game Management’ they say: “We predict that Curlew abundance and population change will be positively associated with the extent of strip burning, through its role in creating variation in vegetation height and as a proxy for for grouse moor management, and consequently predator control.” They go on to say: “We predicted that Curlew abundance and population change would be positively associated with Red Grouse and Pheasant numbers, as a proxy for aspects of game management such as predator control.” Under the section headed ‘Predator populations’, they say: “We predict that Curlew abundance and population change will be negatively associated with both crow and fox occurrence.”
So we can safely conclude that direct predator control works; and that its use is sufficiently rapid in its positive effects for species of conservation interest. It is swift and efficient in its effects.
There are considerable differences between the British landscape and that of many parts of Europe and North America. One of the main differences is that of land ownership. Almost all land in the UK is owned by someone. Whereas much of the wild, undeveloped land in North America (and a good deal of Europe) is de facto owned by the state in the sense that exploitation needs licenses from the state to develop or extract from it in some way. In the UK the ownership of land means that the demand to make that land provide an income is very strong. This means that the land is worked very hard to provide the owner or tenant with an income. As this pattern of ownership has progressed over the centuries, the landscape has been shaped into what we see today – a highly diverse patchwork quilt of woodlands, fields and open spaces on a scale which is much smaller than in many other parts of the world. But it gives us a biological, social, economic, historical and cultural diversity which is unique and very, very special. If the rewilders wish to change all that, then they need an extraordinarily good reason to do so. So far, neither the rewilding movement as a whole, nor Steve Carver, have provided that reason.
References:
[1] Begon M, Townsend CR, Harper JL (2006): Ecology – from Individuals to Ecosystems (Fourth edition). Blackwell Publishing, Oxford. [2] Serrouya R, McLellan BN, van Oort H, Mowat G and Boutin S (2107): Experimental moose reduction lowers wolf density and stops decline of endangered caribou.PeerJ5:e3736;DOI10.7717/peerj3736. [3] Potts GR, (2012): Partridges; Harper Collins, London. [4] Newton I (2017): Farming and Birds. Chapter 13, pp 338-345. Harper Collins, London. [5] Franks SE, Douglas DJT, Gillings S and Pearce-Higgins JW (2017): Environmental correlates of breeding abundance and population change of Eurasian Curlew (Numenius arquata) in Britain. Bird Study, DOI 10.1080/00063657.2017.1359233.David Eyles is a retired livestock farmer with a past life as a quantity surveyor in civil engineering. He now lives in Cornwall, where he indulges his lifelong interests in birds, natural history, and walking. He occasionally finds sanctuary in datasets with long time series, and writes about subjects which attract his attention. His blog is here.
2 thoughts on “Managing Predators & Prey”
Comments are closed.